Sebastian Mueller, MD, PhD
Professor of Medicine,
CAR, University of Heidelberg
VISCERA AG Bauchmedizin Bern
|
Sebastian Mueller, MD, PhD
Professor of Medicine, CAR, University of Heidelberg
VISCERA AG Bauchmedizin Bern
|
|
|
Based on our novel ROS models (see Biochemistry section), we were able to study how these small oxygen derived molecules such as H2O2, hypochlorous acid or others are involved in the communication within or between cells. These studies are important for the understanding of impaired cellular signaling has is seen during inflammation or in cancer tissues. Another interest has been, how these reactive oxygen species (ROS) are actually produced, metabolized and compartmentilized in cells. Below, some previous key findings are briefly listed.
Selected topics
1. Release and metabolsism of H2O2 from liver peroxisomes 2. Hypoxia signaling exploring a novel enzymatic hypoxia system 3. Regulation of iron metabolism by reactive oxygen species (ROS)
5. References hypoxia signaling
1. Release and metabolism of H2O2 from liver peroxisomes
Fig. 1
Fig. 1 shows the release of H2O2 via very small channels (A) in the cytosol (B) of liver cells. The paper has been the first to directly demonstrate that peroxisomes which are ubiquituous and essential cell organelles are not only detoxifiers of H2O2 but can readily release this molecule into the surrounding cytosol where it possibly could serve as second messenger and metablic control.
2. Hypoxia signaling exploring a novel enzymatic hypoxia system
Fig. 2
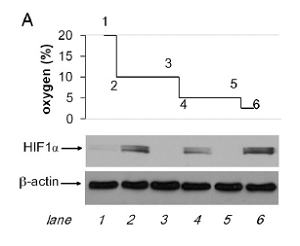
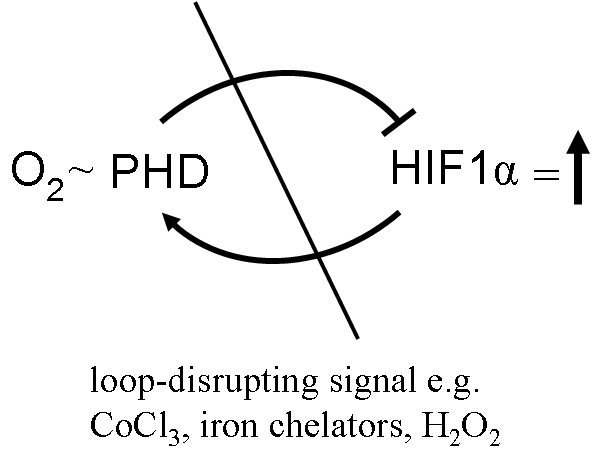
Using the GOX/CAT system (see Biochemistry section) we could show that the important transcription factor hypoxia inducible factor 1alpha (HIF1 alpha) is not responding to oxygen levels but to the decrease of oxygen within a certain time period. Fig. 2 shows that nuclear HIF1alpha (red) is only seen after 2 hours of hypoxia but not after 24 hours although the liver cells are alive and have actually adapted to these low oxygen levels. Fig. 3 shows directly that lowering oxygen by 50% will result in equal expression levels of HIF1 no matter what absolute degree of hypoxia is present. We have interpreted the negative feed back loop of HIF1-degrading prolyl hydroxylases (PHD) and HIF1 (see Fig. 4) as metabolic differential control. Only a disruption of this loop will result in sustained upregulation of HIF1 which will increase the metabolic turnover of tissues (for details see also ref. 27. and 36.).
Fig. 4
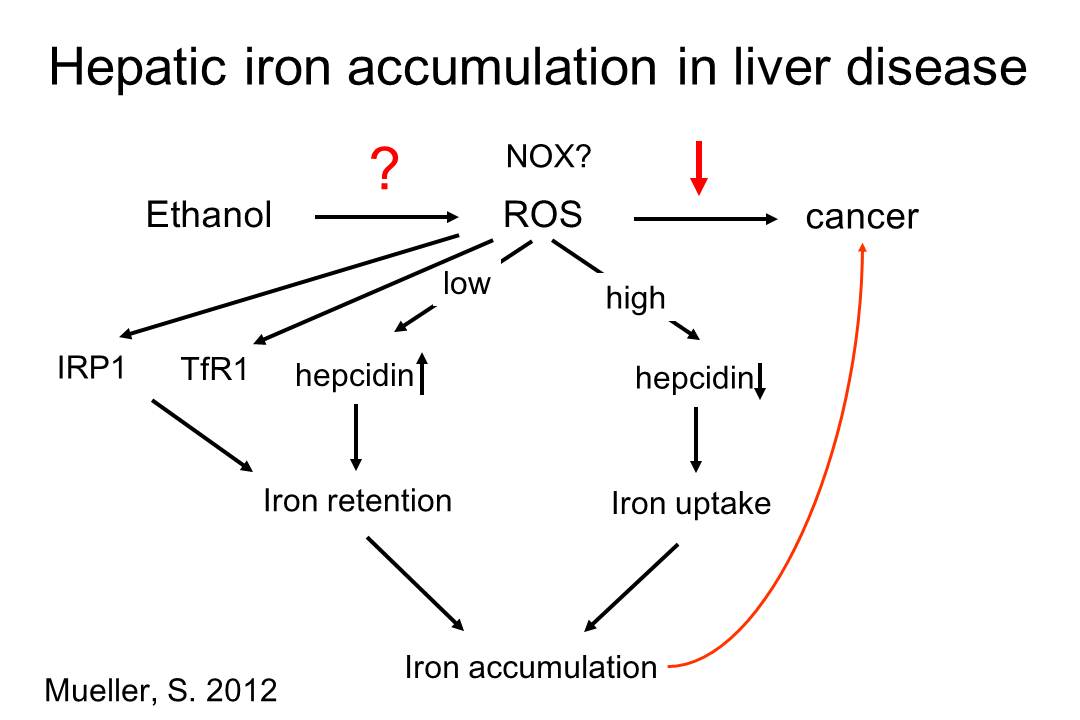
3. Regulation of iron metabolism by reactive oxygen species (ROS) Iron overload is a major risk factor for liver cancer and patients with chronic liver disease or cirrhosis commonly show increased levels of iron in their liver such as alcoholic liver disease or chronica hepatitis C (see Fig. 5 below). The toxicity and carcinogenicity of iron is mainly explained by its high reactivity with reactive oxygen species (ROS) that eventually lead to extremely reactive hydroxyl radicals via the Fenton chemistry. Since the diseased liver shows a high ROS-turnover even mildly increased iron levels can be highly carcinogenic. On the other side, the molecular mechanism of the hepatic iron accumulation in chronic liver diseases are poorly understood but could provide novel targeted approaches to prevent iron-based carcinogenesis in chronic liver disease. Fig. 5
We have a longterm research focus on how ROS regulate key proteins of iron metabolism at the cellular and systemic level. In previous studies we have shown that the central ROS metabolite hydrogen peroxide (H2O2) causes cellular iron accumulation via posttranscriptional (iron regulatory protein 1) and translational (transferrin receptor 1) mechanisms. Recent work from our laboratory indicates that ROS also affect the systemic control of iron. As shown in Fig. 5 above, a better understanding of the mechanisms underlying iron accumulation and toxicity could help in developing novel targeted therapies.
4. References iron studies
61.
CHOP-mediated
hepcidin suppression modulates hepatic iron load.
Mueller K, Sunami Y, Stuetzle M, Güldiken N, Kucukoglu O, Mueller S, Kulaksiz
H, Schwarz P, Strnad P. J Pathol. 2013 Jun 8. doi: 10.1002/path.4221. [Epub
ahead of print] PMID: 23749468 [PubMed - as supplied by publisher] Related
citations
Millonig G, Ganzleben I, Peccerella T, Casanovas G, Brodziak-Jarosz L, Breitkopf-Heinlein K, Dick TP, Seitz HK, Muckenthaler MU, Mueller S. J Biol Chem. 2012 Aug 29. [Epub ahead of print] PMID: 22932892 PubMed - as supplied by publisher] Free Article Related citations
53. Siderophore-mediated
iron trafficking in humans is regulated by iron. Liu Z, Lanford R, Mueller S, Gerhard GS,
Luscieti S, Sanchez M, Devireddy L. J
Mol Med (Berl). 2012
Apr 15. [Epub
ahead of print] PMID: 22527885 [PubMed
- as supplied by publisher] 44. Increased
iron in HCV infection: Collateral damage or antiviral defense? Mueller
S. J
Hepatol. 2010
Aug 22. [Epub ahead of print] No abstract available. PMID:
20850193 [PubMed - as supplied by publisher] Related
citations 43. Heme
Oxygenase-1 and Iron in Liver Inflammation: a Complex Alliance. Immenschuh
S, Baumgart-Vogt E, Mueller S. Curr
Drug Targets.
2010 Aug 13. [Epub ahead of print]PMID:
20704547 [PubMed - as supplied by publisher] Related
citations 42. Heme
oxygenase-1 as a therapeutic target in inflammatory disorders of the
gastrointestinal tract. Vijayan
V, Mueller S, Baumgart-Vogt E, Immenschuh S. World
J Gastroenterol.
2010 Jul 7;16(25):3112-9.PMID:
20593496 [PubMed - in process]Free
PMC Article Free
textRelated
citations 29. In
vitro-targeted gene identification in patients with hepatitis C using a
genome-wide microarray technology.
Hepatology.
2009 Feb;49(2):378-86. PMID:
19177570 [PubMed - indexed for MEDLINE] Related
Articles
25.
Sustained
hydrogen peroxide induces iron uptake by transferrin receptor-1
independent of the iron regulatory protein/iron-responsive element network.
J
Biol Chem. 2007 Jul 13;282(28):20301-8. Epub 2007 May 21. PMID: 17517884 [PubMed
- indexed for MEDLINE] Related
Articles Free
article at journal site
24.
Compartment-dependent
management of H(2)O(2) by peroxisomes.
Free
Radic Biol Med. 2007 Apr 1;42(7):1119-29. Epub 2007 Jan 12. PMID: 17349938
[PubMed - indexed for MEDLINE] Related
Articles
23.
Liver-homing
of purified glucose oxidase: a novel in vivo model of physiological
hepatic oxidative stress (H2O2).
J
Hepatol. 2007 Mar;46(3):482-91. Epub 2006 Nov 17. PMID:
17188390 [PubMed - indexed for MEDLINE] Related
Articles
22. Iron,
HCV, and liver cancer: hard metal setting the pace?
Gastroenterology.
2006 Jun;130(7):2229-34. No abstract available.PMID: 16762645 [PubMed -
indexed for MEDLINE] Related
Articles
21.
Iron
regulatory protein 1 as a sensor of reactive oxygen species.
Biofactors.
2005;24(1-4):171-81. PMID: 16403978 [PubMed - indexed for MEDLINE] Related
Articles
18.
Extracellular
H2O2 and not superoxide determines the compartment-specific activation of
transferrin receptor by iron regulatory protein 1.
Free
Radic Res. 2005 Aug;39(8):817-24. PMID:
16036361 [PubMed - indexed for MEDLINE]
16.
Myeloperoxidase-derived
hypochlorous acid antagonizes the oxidative stress-mediated activation of
iron regulatory protein 1. J
Biol Chem. 2003 Oct 17;278(42):40542-9. Epub 2003
Jul 29. Erratum in: J Biol Chem. 2003 Dec 5;278(49):49662. PMID:
12888561 [PubMed - indexed for MEDLINE] Related
Articles Free
article at journal site
15.
Sensitive
and real-time determination of H2O2 release from intact peroxisomes.
Biochem
J. 2002 May 1;363(Pt 3):483-91. PMID:
11964148 [PubMed - indexed for
MEDLINE] Related
Articles Free
article in PMC | at journal site
14.
Activation
of iron regulatory protein-1 by oxidative stress.
Methods
Enzymol. 2002;348:324-37. PMID: 11885287 [PubMed - indexed for MEDLINE]
12.
IRP1
activation by extracellular oxidative stress in the perfused rat liver.
J
Biol Chem. 2001 Jun 22;276(25):23192-6. Epub 2001 Apr 10. PMID:
11297549 [PubMed - indexed for MEDLINE] Related
Articles Free
article at journal site
Cancer
Res. 2000 Jan 1;60(1):156-63. PMID:
10646868 [PubMed - indexed for MEDLINE] Related
Articles Free
article at journal site
6.
Differences
in the regulation of iron regulatory protein-1 (IRP-1) by extra- and
intracellular oxidative stress.
J
Biol Chem. 1997 Apr 11;272(15):9802-8. PMID:
9092514 [PubMed - indexed for MEDLINE] Related
Articles Free
article at journal site
5. References hypoxia signaling
50. The
effect of low oxygen with and without steady-state hydrogen peroxide on
cytokine gene and protein expression of monocyte-derived macrophages -
biomed 2011.
48.
Investigation
of tumor hypoxia using a two-enzyme system for in vitro generation of
oxygen deficiency. Askoxylakis
V, Millonig G, Wirkner U, Schwager C, Rana S, Altmann A, Haberkorn U,
Debus J, Mueller S, Huber PE. Radiat
Oncol.
2011 Apr 10;6:35. Free PMC Article PMID:21477371[PubMed
- in process] Free
full text Related
citations 27.
Hypoxia-inducible
factor 1a under rapid enzymatic hypoxia: Cells sense decrements of oxygen
but not hypoxia per se. Free
Radic Biol Med. 2009 Jan 15;46(2):182-191. Epub 2008
Nov 1. PMID:
19007879 [PubMed - as supplied by publisher] Related
Articles
45.
Ethanol-mediated
carcinogenesis in the human esophagus implicates CYP2E1 induction and the
generation of carcinogenic DNA-lesions. Millonig
G, Wang Y, Homann N, Bernhardt F, Qin H, Mueller S, Bartsch H, Seitz HK. Int
J Cancer. 2011
Feb 1;128(3):533-40.PMID:
20715111 [PubMed - indexed for MEDLINE]Related
citations 32. Alcoholic
liver disease and hepatitis C: a frequently underestimated combination.
World
J Gastroenterol. 2009 Jul 28;15(28):3462-71. PMID:
19630099 [PubMed - in process] Related
Articles Free
article in PMC | at journal site
31. Ethanol-induced
cytochrome P4502E1 causes carcinogenic etheno-DNA lesions in alcoholic
liver disease. Hepatology.
2009 Aug;50(2):453-461. PMID: 19489076 [PubMed - as supplied by publisher]
Related
Articles
28. Alcohol
and colorectal cancer: the role of alcohol dehydrogenase 1C polymorphism.
Alcohol
Clin Exp Res. 2009 Mar;33(3):551-6. Epub 2008 Dec 19. PMID:
19120062 [PubMed - indexed for MEDLINE] Related
Articles 23.
Liver-homing
of purified glucose oxidase: a novel in vivo model of physiological
hepatic oxidative stress (H2O2).
J
Hepatol. 2007 Mar;46(3):482-91. Epub 2006 Nov 17. PMID:
17188390 [PubMed - indexed for MEDLINE] Related
Articles
Cancer
Res. 2000 Jan 1;60(1):156-63. PMID:
10646868 [PubMed - indexed for MEDLINE] Related
Articles Free
article at journal site
8.
Role
of p53 in aziridinylbenzoquinone-induced p21waf1 expression.
Related
Articles Free
article at journal site
|

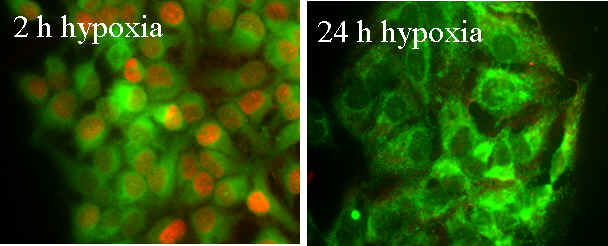 Fig. 3
Fig. 3